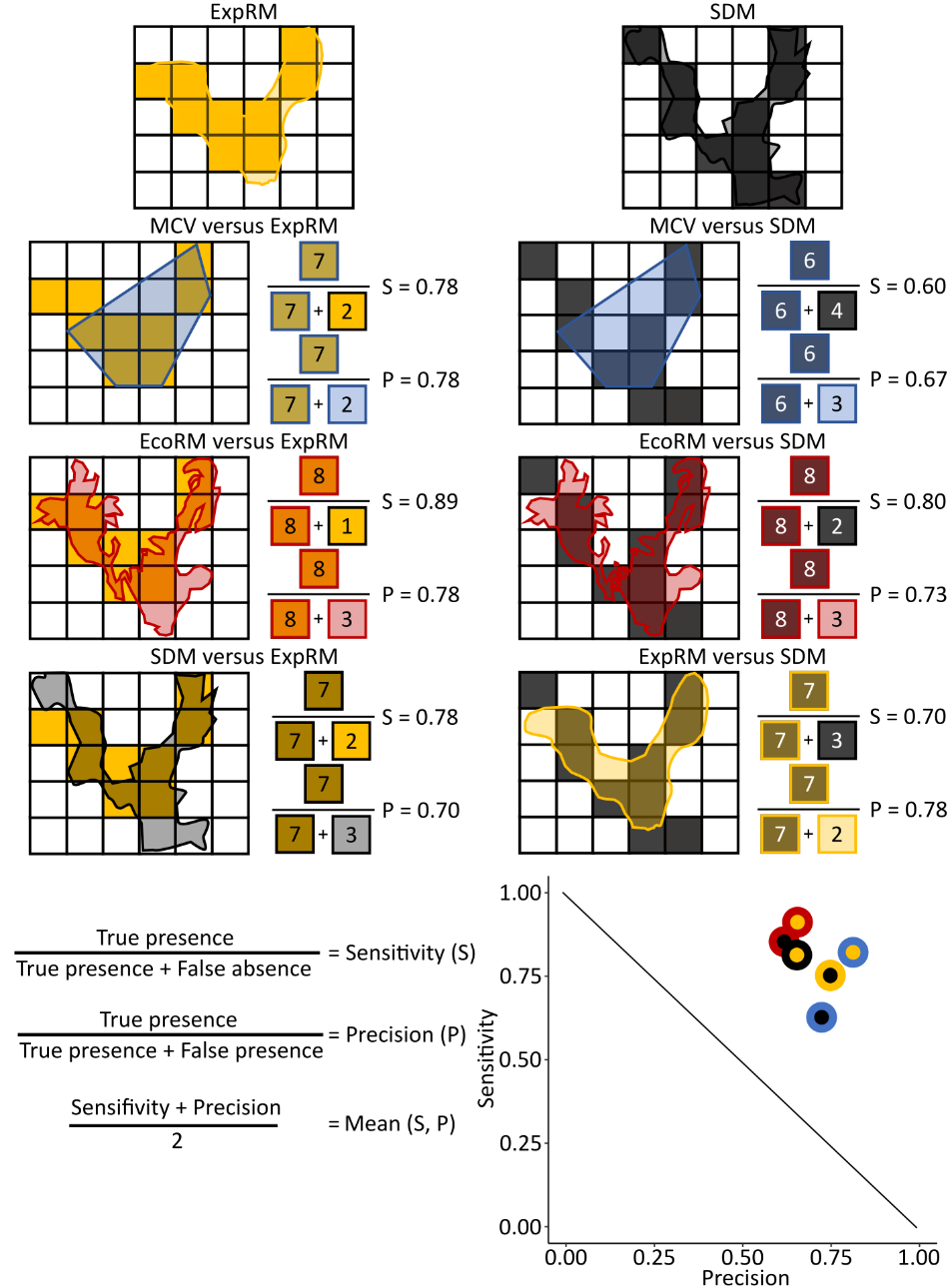
INTRODUCTIONInformation about species’ geographical distribution is central to many ecological and evolutionary questions and underpins effective conservation decision‐making (Meyer et al., 2015; Jetz et al., 2019; Oliver et al., 2021; Jetz et al., 2022). Ideally, distributional data for a species is contiguous in space and time and covers its entire global distribution, at a scale reasonable to inform conservation action and research (Jetz et al., 2019). Expert range maps (ExpRMs) arguably come closest to this standard (Rondinini et al., 2006). These maps of aggregated knowledge and field experience about a species range have been the most frequently used type of distributional data in global-scale biogeographical analyses, biodiversity research, and area-based conservation (Hurlbert & Jetz, 2007; Herkt et al., 2017; Jung et al., 2021). Nevertheless, their limited taxonomic scope critically hampers conclusions about the vast majority of species on our planet (Dauby et al., 2017).The importance of ExpRMs in conservation stems from their use in assessing crucial measures of a species’ threat status by the International Union for the Conservation of Nature [IUCN Red List criterion B; see Schatz (2002)]. The IUCN is the largest provider of ExpRMs, currently holding ranges for approximately 115,000 species (iucn.org retrieved March 11, 2022). Given the lack of monitoring data to assess species range size (i.e., B1 - extent of occurrence) and population density (B2 - area of occupancy), these measures are often estimated using ExpRMs. In addition, changes in these measures over time are used to further track population declines and range contractions. Within IUCN specialists groups, experts draw simplified polygons around occurrence records and then refine and/or extend these polygons based on ecological knowledge of the species in concert with habitat layers (Hawkins et al., 2008; IUCN Standards and Petitions Subcommittee, 2022). Typically, many experts are involved in the generation and evaluation of range maps ensuring that IUCN ExpRMs generally provide a reliably high-quality representation of species’ distributions. Other sources of ExpRMs, especially for plant and invertebrate taxa, include monographies of taxa as well as regional and global field guides (e.g., Scott, 1997; Glassberg, 2017).Over the last few years, an increasing effort has been put into mobilizing range maps from literature sources under metadata standards to not only make them publicly available but also for clarifying decisions of the production process (see birdlife.org or mol.org digitized expert ranges; Marsh et al., 2022). However, due in part to the immense work necessary to produce or digitize each ExpRM, their availability is often limited to more popular or well-studied taxa. Currently, comprehensive and high quality ExpRMs are available for a large proportion of vertebrate species (IUCN.org), whereas they are available only for a few selected plants and invertebrates subgroups and typically limited in their geographical extent (e.g. mol.org/patterns).Species occurrence data, particularly from museum collections and citizen science efforts, have grown rapidly in recent decades. The Global Biodiversity Information Facility hosts occurrence records for 1,723,634 animal and plant species (gbif.org retrieved March 11, 2022). Approaches integrating ExpRMs with increasingly complete, spatially explicit, and readily available occurrence data promise unique advances for incorporating a significant proportion of all species on Earth into large-scale assessments on the status and trend of biodiversity. The IUCN has adopted two alternative integrative approaches to address limitations of transparency and reproducibility of ExpRMs. First, hydro basin layers are used to infer species’ ranges from intersections with observation- and literature-based occurrence records. Although hydro basin-based ranges are limited to species affiliated with lotic (running) waters, this approach vastly improved the availability of baseline distributional data for the assessment of species’ threat status of crabs, crayfishes, shrimps, and Odonata (IUCN.org). Secondly, simple, non-parametric occurrence-based estimates such as Minimum Convex Hulls (MCVs) have been proposed as range estimates to calculate the extent of occurrence and the area of occupancy of species (Dauby et al., 2017; see also ala.org.au). However, both alternatives do not resolve the internal structure of species’ ranges that result from barriers to dispersal, geological differences, and ecological gradients and are therefore likely to significantly overestimate the true species range in many cases. As a result, these range surrogates should be more sensitive (i.e., cover more suitable habitat or potential presences) but less precise (i.e., have a lower occupancy of suitable habitat) than ExpRMs, at least for data-rich species (see Figure 1).We propose the use of terrestrial ecoregions to develop alternative ExpRM surrogates, what we denominate Ecoregional Range Maps (EcoRMs). Ecoregions define the natural extent of areas with similar environmental conditions and distinct ecological communities. Freshwater, marine, and terrestrial ecoregions are established baseline layers used in conservation efforts by the World Wildlife Fund and The Nature Conservancy as well as in assessments of the progress of conservation strategies (e.g., Sayre et al., 2014; Dinerstein et al., 2020). Being based on broad geological and ecological zonation, ecoregions imply a high surrogacy value for species distributions of a broad spectrum of organisms but their congruence with single species distributions and biodiversity patterns has thus far not been evaluated.Here, we statistically compare the sensitivity and precision of MCVs and EcoRMs based on predicted absence-presence information from ExpRMs and SDMs at the species-level as well as congruence in the resulting species richness patterns. We generally expect MCVs to cover areas with a high proportion of true presences (i.e., to have a higher precision), but EcoRMs to cover a higher overall number of species’ true presences (i.e., to have a higher sensitivity). In addition, we investigate the spatial dependence as well as the relationships of sensitivities and precision with the number of underlying occurrence records. With these evaluations, we aim to inform applications about the potential of non-parametric, readily applicable, updateable, and occurrence-based alternatives to ExpRMs and SDMs for boosting the integration of data-poor species into both conservation and ecological research.