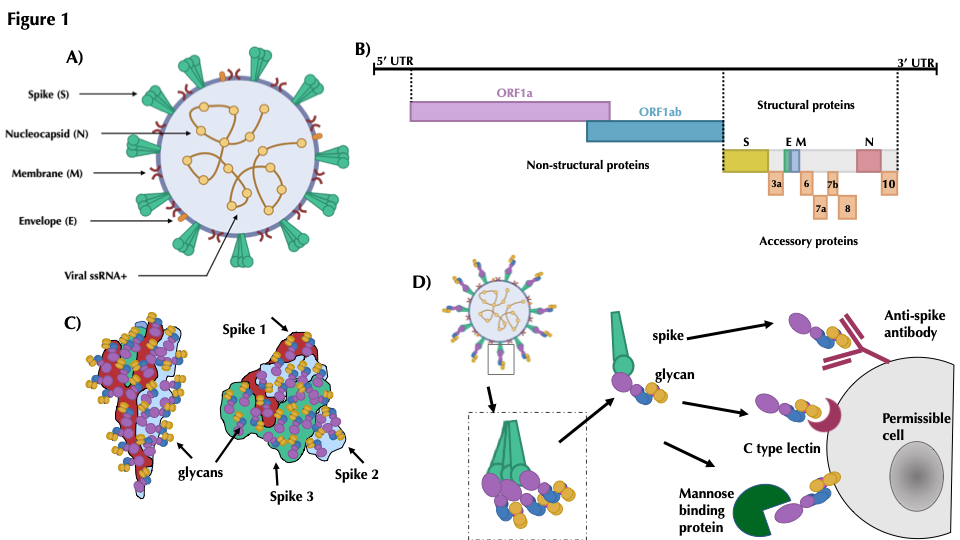
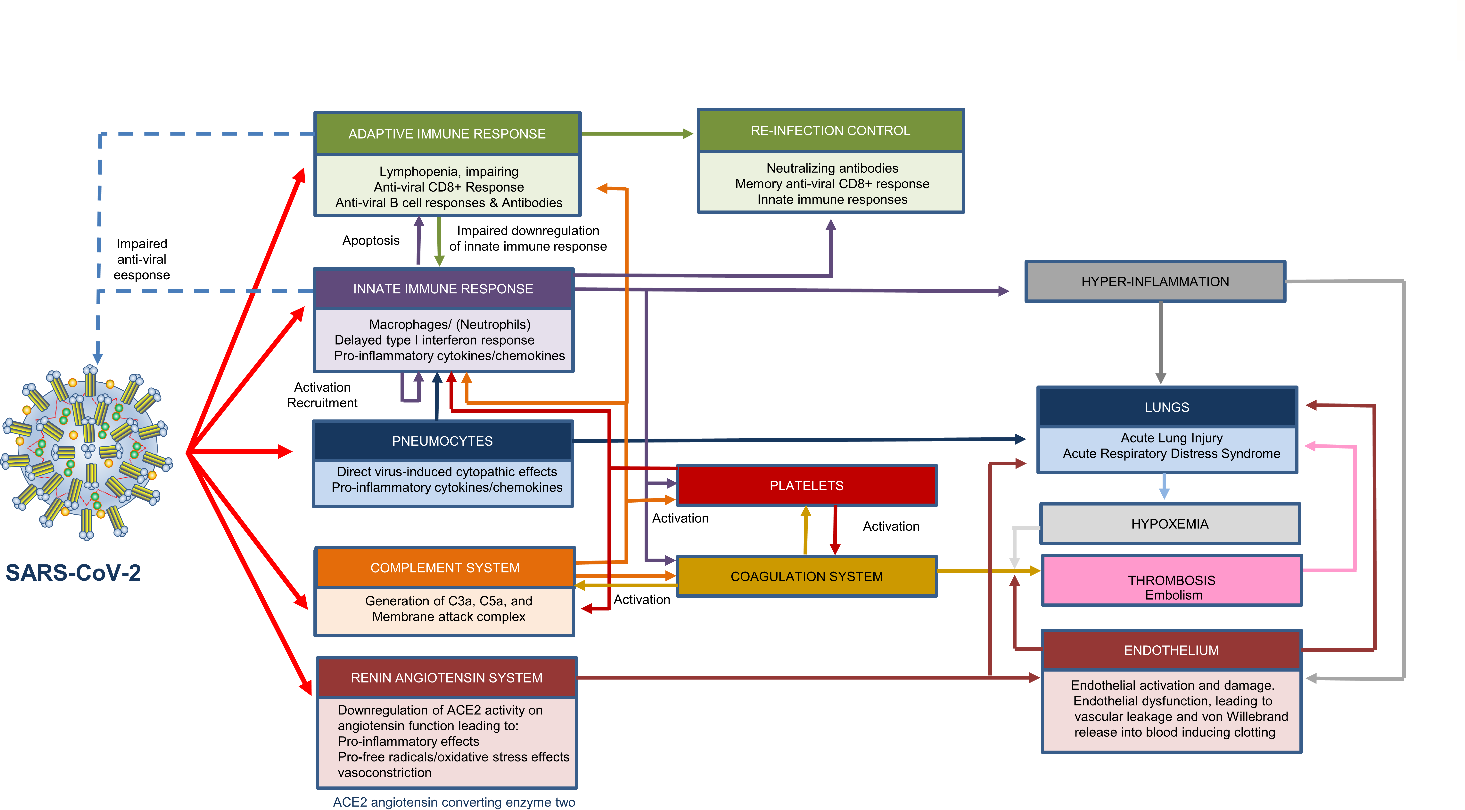
Although most autoimmune diseases are considered to be CD4 T-cell or antibody-mediated, many respond to CD20-depleting antibodies that have limited influence on CD4 and plasma cells. This includes rituximab that is used in cancer, rheumatoid arthritis and off-label in a large number of other autoimmunities, notably multiple sclerosis, where ofatumumab is in late stage development and ocrelizumab is approved for use. Recently, the COVID-19 pandemic created concerns about immunosuppression in autoimmunity, leading to cessation or a delay in immunotherapy treatments. However, based on the known and emerging biology of multiple sclerosis and COVID-19, it was hypothesised that whilst B-cell depletion should not necessarily expose people to severe SARS-CoV-2-related issues, it may inhibit protective immunity following infection and vaccination. As such, drug-induced B-cell subset inhibition that controls multiple sclerosis and other autoimmunities, would not influence innate and CD8 T-cell responses, which are central to SARS-CoV-2 elimination, nor the hyper-coagulation and innate inflammation causing severe morbidity. This is supported clinically, as the majority (mortality rate n=~5/392) of SARS-CoV-2 infected, CD20-depleted people with multiple sclerosis have recovered. However, protective neutralising-antibody and vaccination responses are predicted to be blunted, until naïve B-cells repopulate, based on B-cell repopulation-kinetics and vaccination responses, from published rituximab and unpublished ocrelizumab (NCT00676715, NCT02545868) trial data, shown here. This suggests that it may be possible to undertake dose-interruption to maintain inflammatory disease control in MS and other autoimmune diseases, whilst allowing effective vaccination against SARS-CoV-29, if and when an effective vaccine is available.