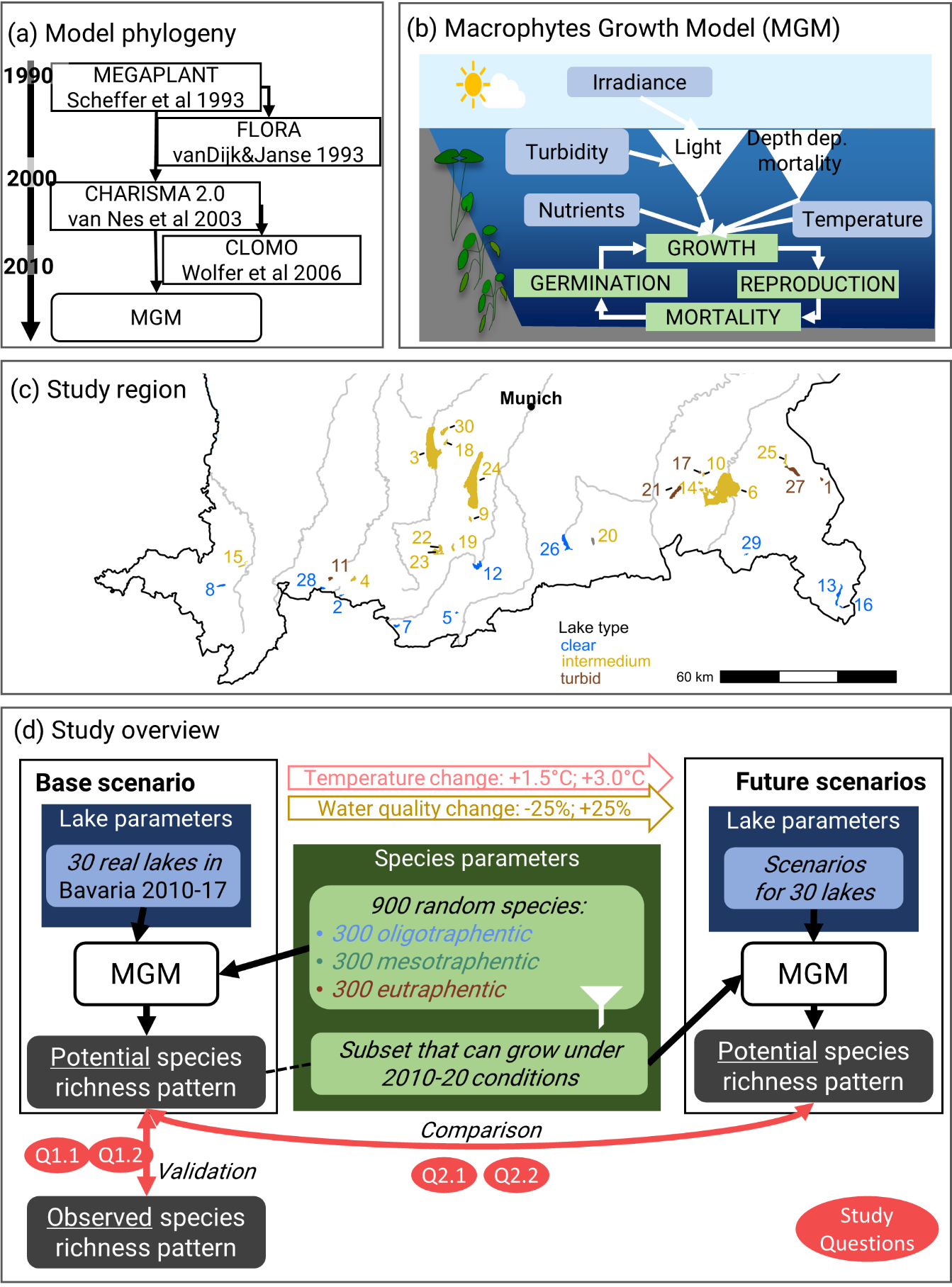
Juliano Sarmento Cabral1, Alma Mendoza-Ponce2,3, André Pinto da Silva4,5, Johannes Oberpriller6, Anne Mimet7, Julia Kieslinger8, Thomas Berger9, Jana Blechschmidt1, Maximilian Brönner8, Alice Classen10, Stefan Fallert1, Florian Hartig6, Christian Hof7, Markus Hoffmann11, Thomas Knoke12, Andreas Krause13, Anne Lewerentz1, Perdita Pohle8, Uta Raeder11, Anja Rammig13, Sarah Redlich10, Sven Rubanschi7, Christian Stetter14, Wolfgang Weisser7, Daniel Vedder1,15,16,17 , Peter H. Verburg18, Damaris Zurell191 Ecosystem Modelling, Center for Computational and Theoretical Biology (CCTB), University of Würzburg, Klara-Oppenheimer-Weg 32, 37074, Würzburg, Germany2 Research Program on Climate Change, Universidad Nacional Autónoma de México, Mexico City, Mexico3 International Institute for Applied Systems Analysis, Laxenburg, Austria4 Department of Ecology and Genetics, Animal Ecology, Evolutionary Biology Centre, Uppsala University, Uppsala, Sweden5 Centre for Ecology, Evolution and Environmental Changes (cE3c), Faculdade de Ciências, Universidade de Lisboa, Lisbon, Portugal6 Theoretical Ecology Lab, University of Regensburg, Universitätsstraße 31, 93053 Regensburg, Germany7 Technical University of Munich, Terrestrial Ecology Research Group, Department of Life Science Systems, School of Life Sciences, 84354 Freising, Germany8 Institute of Geography, Friedrich-Alexander University Erlangen-Nuernberg, Wetterkreuz 15, 91058 Erlangen, Germany9 Land-Use Economics in the Tropics and Subtropics, Hans-Ruthenberg Institute, Hohenheim University, Hohenheim, Germany10 Department of Animal Ecology and Tropical Biology, Biocentre, University of Würzburg, Am Hubland, 97074 Würzburg, Germany11 Technical University of Munich, Limnologische Station Iffeldorf, Chair of Aquatic Systems Biology, Department of Life Science Systems, School of Life Science,Hofmark 1-3, 82393 Iffeldorf, Germany12 Technical University of Munich, Institute of Forest Management, Department of Life Science Systems, School of Life Sciences, 58354 Freising, Germany13 Technical University of Munich, Land Surface-Atmosphere Interactions, Department of Life Science Systems, School of Life Sciences, 85354 Freising, Germany14 Agricultural Production and Resource Economics, School of Life Sciences, Technical University of Munich, 84354 Freising, Germany15 Helmholtz Center for Environmental Research - UFZ, Department of Ecosystem Services, Permoserstr. 15, 04318 Leipzig, Germany16 Institute of Biodiversity, Friedrich Schiller University Jena, Dornburger Straße 159, 07743 Jena, Germany17 German Centre for Integrative Biodiversity Research (iDiv) Halle-Jena-Leipzig, Puschstr. 4, 04103 Leipzig, Germany18 Institute for Environmental Studies, VU University Amsterdam, De Boelelaan 1111, 1081 HV Amsterdam, The Netherlands19 Ecology & Macroecology, Inst. for Biochemistry and Biology, University of Potsdam, Am Neuen Palais 10, 14469 Potsdam, GermanyArticle type: review/perspective