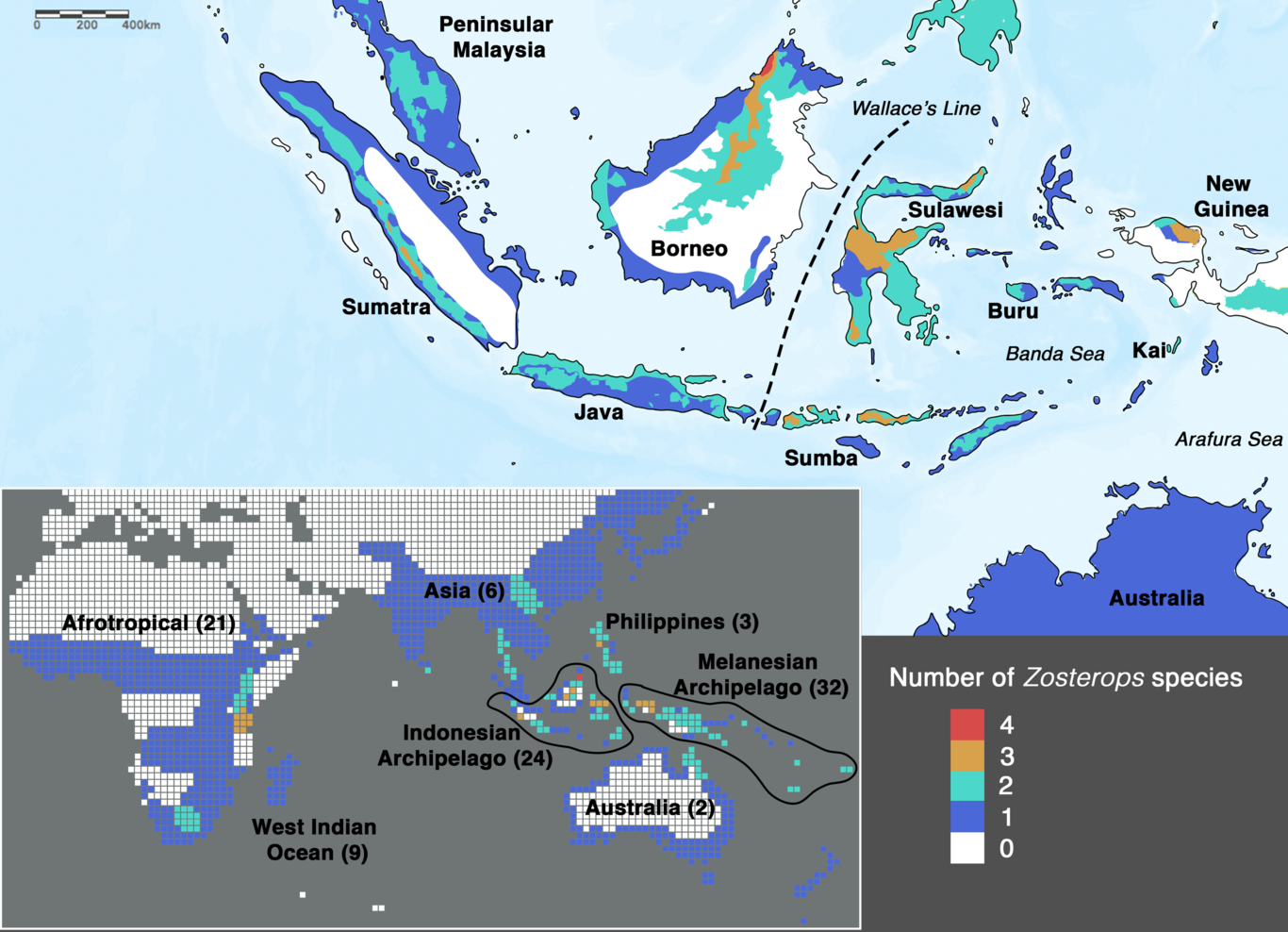
Chyi Yin Gwee, Kritika M. Garg, Balaji Chattopadhyay, Keren R. Sadanandan, Dewi M. Prawiradilaga, Martin Irestedt, Fumin Lei, Luke M. Bloch, Jessica G.H. Lee, Mohammad Irham, Tri Haryoko, Malcolm C.K. Soh, Kelvin S.-H. Peh, Karen M.C. Rowe, Teuku Reza Ferasyi, Shaoyuan Wu, Guinevere O.U. Wogan, Rauri C.K. Bowie, Frank E. RheindtINTRODUCTIONArchipelagoes are ideal settings for unravelling complex evolutionary patterns as they constitute natural laboratories for the study of factors contributing to speciation (MacArthur and Wilson 2001; Whittaker and Fernández-Palacios 2007; Lohman et al. 2011). Among vertebrate groups that occur across archipelagoes, island radiations of birds are most well-studied (Lerner et al. 2011; Lamichhaney et al. 2015). These avian models display a great deal of variability in their diversification rates across islands, which are fundamentally linked to species’ capability to disperse over water (Diamond et al. 1976). In particular, the so-called ‘great speciators’, first characterized by Diamond et al. (1976), stand out from all other birds based on their paradoxical ability to disperse widely and colonize entire archipelagoes while, at the same time, diversifying into multiple daughter species in spite of a continuing potential for overwater gene flow (Cai et al. 2020).One of the few classical examples of ‘great speciators’ identified by Diamond et al. (1976) is the songbird genus Zosterops , or white-eyes, which are excellent dispersers capable of differentiating rapidly from source populations (Clegg et al. 2002; Moyle et al. 2009). The genus Zosterops comprises c. 100 species that have radiated across the Old World and Oceania within the past 1 to 3.5 million years, reflecting perhaps the fastest diversification rate of any vertebrate clade (Warren et al. 2006; Moyle et al. 2009; Leroy et al. 2019; Cai et al. 2020). An overwhelming proportion (more than 70%) of these species occurs exclusively in archipelagoes distributed across the Atlantic, Indian and Pacific Oceans (Fig. 1). As such, the remarkable radiation of white-eyes serves as an ideal model system with which to explore island biogeography theory (Diamond et al. 1976; Moyle et al. 2009).In order to achieve an understanding of the underlying processes driving the white-eye radiation, its phylogeny first needs to be resolved to provide a reliable backbone for hypothesis testing. In Zosterops , however, traditional methods that rely on morphological tools to infer how species are related to one another have proven to be unreliable, as plumage features of ecologically distinct and geographically disjunctZosterops species are often indistinguishable (Mees 1957; Mayr 1965). Although a more recent application of genetic methods has helped disentangle the white-eye radiation to some extent, most studies have concentrated on Melanesian and Indian Ocean members of the genus (Slikas et al. 2000; Warren et al. 2006; Moyle et al. 2009; Cox et al. 2014; Linck et al. 2016; Wickramasinghe et al. 2017; Manthey et al. 2020). There continues to be a dearth of knowledge on this radiation across the core of its Asian distribution due to limited sampling and lack of genetic data. In particular, it is crucial to unravel the phylogenetic affinities of white-eyes distributed across the Indonesian archipelago, which – comprising more than 17,000 islands – is the largest archipelago in the world and harbors about 20 endemic Zosteropsspecies (Fig. 1), including two undescribed species that were discovered in the last two decades (Eaton et al. 2016; O’Connell et al. 2019). The high density of Zosterops species across the Indonesian archipelago hints at the possible importance of this region in white-eye evolution.Apart from incomplete geographic sampling, the lack of resolution of the white-eye radiation has largely been a consequence of sparse genomic sampling: most phylogenetic studies of white-eyes have been restricted to one or a few genetic markers, resulting in trees that are plagued by unresolved polytomies, hampering useful evolutionary inference (Slikas et al. 2000; Warren et al. 2006; Moyle et al. 2009; Oatley et al. 2012; Nyári and Joseph 2013; Cox et al. 2014; Husemann et al. 2016; Linck et al. 2016; Round et al. 2017; Wickramasinghe et al. 2017; Shakya et al. 2018; Cai et al. 2019; Lim et al. 2019; O’Connell et al. 2019). Disentangling relationships within rapid and recent radiations, such as white-eyes, requires overcoming the challenges of heterogenous gene trees due to biological factors such as incomplete lineage sorting (Edwards et al. 2005; Song et al. 2012). The multispecies coalescent (MSC) model offers a promising avenue to overcoming gene tree discordance by allowing the evolutionary histories of each locus to be inferred independently (Song et al. 2012; Liu et al. 2015).An equally debilitating source of bias in reconstructing the phylogeny of rapid and recent radiations such as white-eyes is the potential for member species to engage in secondary gene flow or genetic introgression long after a speciation event has occurred (Rheindt and Edwards 2011; Edwards et al. 2016). Such introgression will be reflected in the phylogenetic signal of a varying proportion of loci, thereby leading astray efforts to search for the true species tree. Multiple analytical approaches have been devised to account for secondary gene flow, such as through tree-based analysis as implemented in Phylogeographic Inference using Approximate Likelihoods (PHRAPL) (Jackson et al. 2017a; Jackson et al. 2017b), or through SNP-based analysis like the ABBA-BABA test, which detects an excess of shared derived alleles between populations (Green et al. 2010; Patterson et al. 2012).In this study, we acquired historical and modern DNA samples from museums and recent material from our own fieldwork at remote locations to achieve a wide geographic representation of white-eyes (Table S1). We designed RNA probes using Z. lateralis (Cornetti et al. 2015) as a reference genome, targeting 832 loci at high coverage to overcome the limitations of missing data expected from degraded DNA of historical samples, thereby recovering a comparable set of loci across both historical and fresh samples (Templeton et al. 2013). Our target capture methodology addresses the recalcitrant persistence of unresolved polytomies in the phylogeny of this rapid radiation by making use of a large set of loci (Cai et al. 2019). We employed three different coalescent species tree methods to assess topological incongruence across tree-building approaches (Liu et al. 2009; Liu et al. 2010; Vachaspati and Warnow 2015). Recognizing that gene flow is commonly observed in recent radiations, we conducted PHRAPL (Jackson et al. 2017b) analysis and performed ABBA-BABA tests (Patterson et al. 2012) to assess introgression between closely related species with incongruent topologies and ultimately elucidate the likely evolutionary history of this complex radiation.