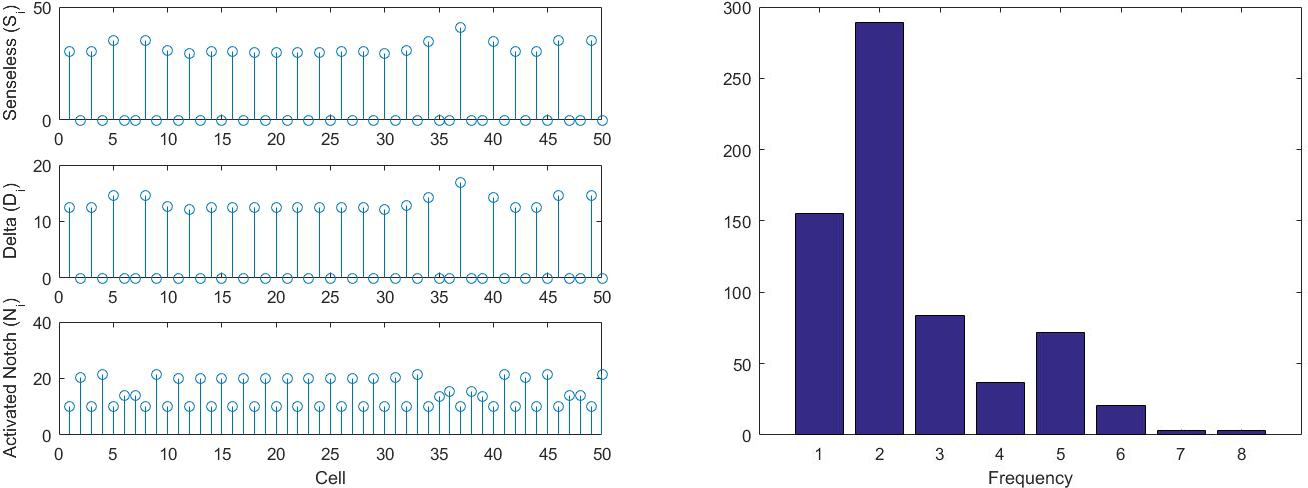
INTRODUCTION Multicellular organisms construct complex morphological structures in an extremely reproducible manner. Reproducibility, in part, is achieved by expressing the right developmental gene in the right pattern. Even so, not all patterns are predetermined. Some patterns arise by cell-cell interaction in a stochastic manner. One such cellular interaction mode is termed ‘lateral inhibition’. In this mode, each cell interacts only with its neighboring cells. Each cell expresses an inhibitory surface molecule called _Delta_ and its cognate receptor molecule _Notch_. _Delta_ expression is driven by a pro-differentiation transcription factor called _Senseless_. Conversely, _Notch_ activation, by the _Delta_ molecule of a neighboring cell, leads to down-regulation of _Senseless_. Thus cells compete to increase their _Senseless_ expression while shutting down _Senseless_ production in their immediate neighbors. Within a competent group of cells, the individual cell with the highest _Senseless_ expression is selected to differentiate into a sensory organ ( and references therein). Lateral inhibition has been used to justify the emergence of periodic patterns of sensory organs that arise in many contexts. One such context is the fruit fly anterior wing margin. Here, around 20-22 chemosensory organs arise in a robust periodic pattern such that every 5th cell in a 1-D row is a sensory organ. The aim of this project is to determine if the dynamics of lateral inhibition can indeed explain the emergence of periodic patterns, and specifically the periodicity of every 5th cell differentiating observed in the fly wing. And if so, how might the period be controlled? PRELIMINARY MODEL We consider a 1D row of cells representing cells on the fruit fly anterior wing margin. We model the dynamics of _Senseless_ Sj, _Delta_ Dj, and activated _Notch_ Nj, where j is the cell index. An externally-imposed inductance signal is given by I, which is taken to be constant. {dT} &= -k_1 S_i + k_2 I F(N_j), \qquad &F(N) \in [0,1], \ F'(N) < 0,\\ {dT} &= -k_3 D_i + k_4 G(S_j), \qquad &G(S) \in [0,\infty), \ G'(S) > 0, \\ {dT} &= -k_5 N_i + k_3 _j, \qquad &_j = (D_{j-1} + D_{j+1})/2. This model is based on the following assumptions: 1. _Senseless_ Sj is removed from the system at rate k₁ due to degradation. 2. _Senseless_ Sj is transcribed at a rate that is proportional to the inductance signal I and a decreasing function of the activated _Notch_ F(Nj). It is believed that _Senseless_ transcription is highly sensitive to the amount of bound _Notch_, so it is plausible that F is sigmoidal (i.e. F(N)=1/(1 + ea(N − Nc))), where increasing a increases sensitivity, and Nc is the amount of _Notch_ where _Senseless_ transcription starts to become affected. 3. _Delta_ is lost at a rate k₃ due to _Notch_ binding with adjacent cells. We assume that the amount of unbound _Notch_ on adjacent cells is at equilibrium (i.e., when _Notch_ becomes bound, it is immediately replaced). 4. _Delta_ is transcribed at a rate that is proportional to an increasing function of the amount of _Senseless_, G(Sj). 5. Activated _Notch_ is lost at a rate k₅ due to degradation, which may occur after the regulatory subunit performs its action in the cell nucleus. 6. _Notch_ is activated at a rate proportional to the average of _Delta_ in the neighboring cells, $_j = (D_{j-1} + D_{j+1})/2$. The rate parameter is chosen to be k₃ so that, over a given interval of time, the amount of _Delta_ lost by all cells due to _Notch_ binding is equal to the amount activated _Notch_ that is created. ANALYSIS In order to determine analytically which periodicities are most linearly-unstable, we conduct a Fourier analysis. For this we take the Fourier ansatz (S_j, D_j, N_j)^T = (_h, _h, _h)^T + (_j, _j, _j)^T e^{\omega t} e^{i k j}, where $(_h, _h, _h)^T$ is a spatially homogeneous steady state solution to the system and k is the Fourier wavenumber. Plugging this ansatz in to the system gives the eigenvalue problem $\omega = J $ at linear order, where J is the Jacobian matrix of the system linearized about the homogeneous steady state. By solving for the largest eigenvalue of J, we find that modes of k = π/2, π, 3π/2, ... are most unstable, which correspond to periodicities of two and four. In numerical simulations we have observed the 2-periodic pattern but not the 4-periodic pattern. Also it appears that a 5-periodic pattern cannot be most unstable in any parameter regime. MODIFIED MODEL From our analysis, it appears that the preliminary model does not easily account for the higher periodicities that are observed in the anterior wing margin. It is possible that the preliminary model does not capture a key biological mechanism that is required to generate these periodicities. It has been experimentally observed in _C. elegans_ that some _Delta_ homologs do not have a transmembrane domain and thus can be secreted by cells, so that they are free floating and are not bound to any membrane. Further, these secreted ligands are functionally equivalent to the membrane bound homologs . We capture this phenomenon by including non-local terms where some fraction of _Delta_ molecules can be secreted. These are then free to bind to non-adjacent cells’ receptors and inhibit _Senseless_ production in cells further away. We modify two terms in the preliminary model: one in the _Notch_ equation to account for the non-local _Delta_ binding and another in the _Delta_ equation to account for the loss of _Delta_ due to this binding. It is not clear what non-local kernel should be used to describe the likelihood of secreted _Delta_ binding to cells of some distance away, since it is not biologically known how secreted _Delta_ propagates. We restricted our numerical study to symmetric kernels which decay away from the origin. In our numerical simulations, we observed that fast-decaying kernels, such as ones that go as 1/r², reproduce the two-cell period seen in the preliminary model (Figure 1). We believe that the effect of non-locality is minimal in this case. However, for kernels which decay more slowly, such as kernels which decay linearly, higher periodicities of 4, 5, and 6 appear in the numerical simulations (Figure 2). CONCLUSION We conclude that our idealization of lateral inhibition is unable to produce the periodicities that are observed in the fruit fly anterior wing margin, and hypothesize that an additional mechanism or molecule is needed to generate a 5-periodic pattern. Since the 2-periodic pattern appears most robustly in both our original model formulation and a plausible modification, we wonder whether it is possible that the initial pattern is in fact every other cell and later events (such as cell division) produce the higher period we see in the adult. Alternatively, our model modifications suggest that a secreted ligand can generate a 5-periodicity, and we suggest further investigation of _Delta_ secretion in the wing margin. There is also evidence for other secreted inhibitors acting during sensory organ specification such as _Scabrous_, a short range inhibitor . Similarly _Fringe_ is expressed in the wing margin and is capable of modifying _Delta_ potency for _Notch_ activation . Although _Fringe_ is a secreted molecule, it is unclear if its effects on _Notch_ signaling are mediated by extracellular _Fringe_ or are cell-autonomous. Nevertheless, it would be worthwhile to investigate if _Scabrous_ or _Fringe_ have a role in periodic pattern formation.